G-белки. Понятие и классификация. G-белок сопряженные рецепторы. Механизм работы рецепторов сопряженных с G-белком G белки строение и функции
Среди органических веществ белки , или протеины , - самые многочисленные, наиболее разнообразные и имеющие первостепенное значение биополимеры. На их долю приходится 50 - 80% сухой массы клетки.
Молекулы белков имеют большие размеры, поэтому их называют макромолекулами . Кроме углерода , кислорода , водорода и азота , в состав белков могут входить сера, фосфор и железо. Белки отличаются друг от друга числом (от ста до нескольких тысяч), составом и последовательностью мономеров. Мономерами белков являются аминокислоты (рис. 1)
Бесконечное разнообразие белков создается за счет различного сочетания всего 20 аминокислот. Каждая аминокислота имеет свое название, особое строение и свойства. Их общую формулу можно представить в следующем виде:
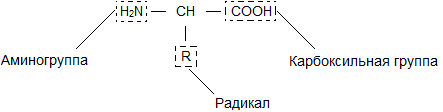
Молекула аминокислоты состоит из двух одинаковых для всех аминокислот частей, одна из которых является аминогруппой (-NH 2 ) с основными свойствами, другая - карбоксильной группой (-COOH ) с кислотными свойствами. Часть молекулы, называемая радикалом (R ), у разных аминокислот имеет различное строение. Наличие в одной молекуле аминокислоты основной и кислотной групп обусловливает их высокую реакционную способность. через эти группы происходит соединение аминокислот при образовании белка. При этом возникает молекула воды, а освободившиеся электроны образуют пептидную связь. Поэтому белки называют полипептидами .
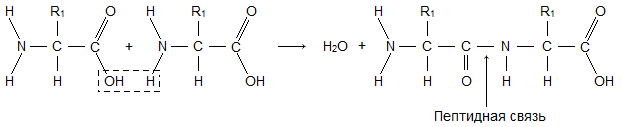
Молекулы белков могут иметь различные пространственные конфигурации, и в их строении различают четыре уровня структурной организации.
Последовательность аминокислот в составе полипептидной цепи представляет первичную структуру
белка. Она уникальна для любого белка и определяет его форму, свойства и функции.
Большинство белков имеют вид спирали в результате образования водородных связей между -CO-
и -NH-
группами разных аминокислотных остатков полипептидной цепи. Водородные связи малопрочные, но в комплексе они обеспечивают довольно прочную структуру. Эта спираль - вторичная структура
белка.
Третичная структура - трехмерная пространственная «упаковка» полипептидной цепи. В результате возникает причудливая, но для каждого белка специфическая конфигурация - глобула . Прочность третичной структуры обеспечивается разнообразными связями, возникающими между радикалами аминокислот.
Четвертичная структура
характерна не для всех белков. Она возникает в результате соединения нескольких макромолекул с третичной структурой в сложный комплекс. Например, гемоглобин крови человека представляет комплекс из четырех макромолекул белка.
Такая сложность структуры белковых молекул связана с разнообразием функций, свойственных этим биополимерам.
Нарушение природной структуры белка называют денатурацией
. Она может происходить под воздействием температуры, химических веществ, лучистой энергии и других факторов. При слабом воздействии распадается только четвертичная структура, при более сильном - третичная, а затем - вторичная, и белок остается в виде полипептидной цепи.
Этот процесс частично обратим: если не нарушена первичная структура, то денатурированный белок способен восстанавливать свою структуру. Отсюда следует, что все особенность строение макромолекулы белка определяются его первичной структурой.
Кроме простых белков , состоящих только из аминокислот, есть еще и сложные белки
(С protein coupled receptors, GPCR), передают сигнал от первичных месенджеров к внутриклеточным мишеням с помощью каскада GPCR-^-G-белок^-эффекторный белок. Первичными сигналами для этих рецепторов служат самые разнообразные молекулы, например, низкомолекулярные гормоны и нейротрансмиттеры (такие, как адреналин, норадреналин, ацетилхолин, серотонин, гистамин), опиоиды, гормоны пептидной и белковой природы (адренокортико- тропин, сомаостатин, вазопрессин, ангиотензин, гонадотропин, эпидермальный фактор роста), некоторые нейропептиды.
В этот же ряд попадают множество химических сигналов, воспринимаемых обонятельными и вкусовыми сенсорными клетками, и свет, рецептором для которого служит пигмент зрительных или фоторецепторных клеток родопсин.
Следует учесть, что один и тот же первичный сигнал может инициировать передачу сигнала через несколько (иногда более 10) разных GPCR, так что, если число внешних сигналов для GPCR составляет несколько десятков, то самих таких рецепторов известно более 200.
При всем их разнообразии GPCR представляют собой мономерные интегральные мембранные белки, полипептидная цепь которых семь раз пересекает клеточную мембрану. Во всех случаях участок рецептора, ответственный за взаимодействие с первичным сигналом, локализован с внешней стороны мембраны, а участок, контактирующий с G-белком - на её цитоплазматической стороне.
Следующий за рецептором компонент каскада передачи сигнала с участием GPCR представлен G-белком. Найдено около 20 различных G-белков, среди них прежде всего нужно упомянуть G s и С;, которые соответственно стимулируют и ингибируют аденилатциклазу; G q , активирующий фосфолипазу С; G-белки сенсорных клеток: фоторецепторных - G t (трансдуцин), обонятельных - G o if и вкусовых - G g .
С-белки - это гетеротримеры, которые состоят из субъединиц трёх типов: ct, (S и у, но в естественных условиях последние две субъединицы функционируют как единый Ру-комплекс. Важнейшая характеристика С-белков - присутствие на их а-субъединице центра связывания гуани- ловых нуклеотидов: ГДФ и ГТФ (рисунки 139, 145). Если с С-белком связан ГТФ, то это соответствует его активированному состоянию. Если в нуклеотидсвязывающем центре присутствует ГДФ, то эта форма соответствует неактивному состоянию белка (рисунок 79).
Центральное событие при передаче сигнала от рецептора, на который подействовал первичный сигнал, к G-белку состоит в том, что активированный рецептор катализирует обмен ГДФ, связанного с G-белком, на присутствующий в среде ГТФ. Такой ГДФ/ГТФ-обмен на G-белке сопровождается диссоциацией тримерной молекулы G-белка на две функциональные субъединицы: а-субъединицу, содержащую ГТФ, и Ру-комп- лекс (рисунки 139, 145).
Далее одна из этих функциональных субъединиц, какая именно - зависит от типа сигнальной системы, взаимодействует с эффекторным белком, представленным ферментом или ионным каналом. Как следствие их каталитическая активность или ионная проводимость соответственно меняется, что, в свою очередь, приводит к изменению цитоплазматической концентрации вторичного месенджера (или иона) и, в конечном счёте, инициирует тот или иной клеточный ответ.
Эффекторными белками в сигнальных системах типа GPCR-^э G-белок^ээффекторный белок могут быть аденилатциклаза, катализирующая синтез цАМФ из АТФ; фосфолипаза С, гидролизующая фосфати- дилинозит с образованием ДАТ и 1Рз; фосфодиэстераза, расщепляющая цГМФ до ГМФ; некоторые типы калиевых и кальциевых каналов.
Важно, что при передаче сигнала в каскаде рецептора G-белок^ээффекторный белок исходный внешний сигнал может многократно усиливаться (амплифицироватъся}. Это происходит благодаря тому, что одна молекула рецептора за время пребывания в активированном состоянии (R*) успевает перевести в активированную форму (G*) несколько молекул G-белка.
Например, в зрительном каскаде родопсин^С^эцГМФ-фосфодиэстераза на каждую молекулу R* может образоваться несколько сотен или даже тысяч молекул G t *, а это означает, что на первой стадии каскада 7?*-»G* коэффициент усиления внешнего сигнала составляет 10 2 -10 3 . Хотя на следующей стадии каскада (С*^ээффекторный белок} каждая молекула G* взаимодействует только с одной молекулой эффекторного белка, сигнал здесь также амплифицируется, поскольку на каждую молекулу G* и, соответственно, активированного эффекторного белка в цитоплазме появляется (или исчезает) множество молекул вторичного месенджера. Так, в зрительном каскаде на второй его стадии одна молекула активированной цГМФ-фосфодиэстеразы способна расщепить в секунду до 3000 молекул цГМФ, служащего в фоторецепторных клетках вторичным месенджером.
Общий коэффициент усиления каскада равен произведению коэффициентов усиления на всех стадиях каскада. Коэффициент амплификации сигнала при его прохождении через каскад может достигать весьма высоких значений: в зрительных клетках это величина порядка 10 5 -10 6 .
Прекращение действия внешнего стимула сопровождается выключением всех компонентов сигнальной системы. На уровне рецепторов это достигается, во-первых, в результате диссоциации первичного месенджера из комплекса с GPCR, во-вторых, путем фосфорилирования рецепторов под действием специальных протеин-киназ и последующего связывания с модифицированным рецептором специального белка (например, Р-аррес- тина).
G-белки обладают ГТФазной активностью, то есть способностью гидролизовать связанный с ними ГТФ до ГДФ, что обеспечивает их самовыключение, то есть переход G-ГТФ^э G-ГДФ. Поскольку состояние активации эффекторного белка (включён-выключен) прямо зависит от состояния G-белка, то этот переход означает также выключение эффекторного белка, а, следовательно, прекращение синтеза (гидролиза) вторичного месенджера или закрывание ионного канала.
И, наконец, чтобы переход клетки к исходному (до действия внешнего стимула) состоянию завершился, специальные механизмы восстанавливают исходный уровень вторичного месенджера или катиона в её цитоплазме. Например, цАМФ, цитоплазматическая концентрация которого повышается при передаче сигнала в каскаде Р-адренорецептора С 5 -белок^аденилатциклаза, гидролизуется затем цАМФ-фосфодиэсте- разой до нециклического (линейного) АМФ, который свойствами вторичного месенджера не обладает.
G-белки (ГТФ-связывающие белки) - универсальные посредники при передаче сигналов от рецепторов к ферментам клеточной мембраны, катализирующим образование вторичных посредников гормонального сигнала. G-белки - олигомеры, состоящие из α, β и γ-субъединиц. Состав димеров βγ незначительно различаются в разных тканях, но в пределах одной клетки все G-белки, как правило, имеют одинаковый комплект βγ-субъединиц. Поэтому G-белки принято различать по их α-субъединицам.. Выявлено 16 генов, кодирующих различные α-субъединицы G-белков. Некоторые из генов имеют более одного белка, вследствие альтернативного сплайсинга РНК.
Каждая α-субъединица в составе G -белка имеет специфические центры:
Связывания ГТФ или ГДФ;
Взаимодействия с рецептором;
Связывания с βγ-субъединицами;
Фосфорилирования под действием протеинкиназы С;
Взаимодействия с ферментом аденилатциклазой или фосфолипазой С.
В структуре G -белков отсутствуют α-спиральные, пронизывающие мембрану домены. G -белки относят к группе «заякоренных» белков (рис. 5-34).
Рис. 5-34. Положение G-белков в мембране. Для ассоциации G-белков важно ацилирование α-протомеров алифатическими радикалами жирных кислот, миристиновой кислоты (М) или изопреновой. γ-Субъединица G-белка имеет геранил-геранильную группу (Г), связанную тиоэфирной связью с остатком цистеина С-конца.
Регуляция активности G -белков
Различают неактивную форму G -белка - комплекс αβγ-ГДФ и активированную форму αβγ -ГТФ. Активация G-белка происходит при взаимодействии с комплексом активатор-рецептор, изменение конформации G-белка снижает сродство α-субъединицы к молекуле ГДФ и увеличивает к ГТФ. Замена ГДФ на ГТФ в активном центре G-белка нарушает комплементарность между α-ГТФ и βγ-субъединицами. Рецептор, связанный с сигнальной молекулой, может активировать большое количество молекул G-белка, таким образом обеспечивая усиление внеклеточного сигнала на этом этапе (рис. 5-35).

Рис. 5-35. Цикл функционирования G-белка. R s - рецептор; Г - гормон; АЦ - аденилатциклаза.
Активированная α-субъединица G-белка (α-ГТФ) взаимодействует со специфическим белком клеточной мембраны и изменяет его активность. Такими белками могут быть ферменты аденилатциклаза, фосфолипаза С, фосфодиэстераза цГМФ, Na + -каналы, К + -каналы.
Следующий этап цикла функционирования G-белка - дефосфорилирование ГТФ, связанного с α-субъединицей, причём фермент, катализирующий эту реакцию, - сама α-субъединица.
Дефосфорилирование приводит к образованию комплекса α-ГДФ, который не комплементарен специфическому белку мембраны (например, аденилатциклазе), но имеет высокое сродство к βγ-протомерам. G-белок возвращается к неактивной форме - αβγ-ГДФ. При последующей активации рецептора и замене молекулы ГДФ на ГТФ цикл повторяется снова. Таким образом, α-субъединицы G-белков совершают челночное движение, перенося стимулирующий или ингибирующий сигнал от рецептора, который активирован первичным посредником (например, гормоном), на фермент, катализирующий образование вторичного посредника.
Некоторые формы протеинкиназ могут фосфорилировать α-субъединицы G-белков. Фосфорилированная α-субъединица не комплементарна специфическому белку мембраны, например, аденилатциклазе или фосфолипазе С, поэтому не может участвовать в передаче сигнала.
Г. аденилатциклаза
Фермент аденилатциклаза, катализирующий превращение АТФ в цАМФ (рис. 5-36), - ключевой фермент аденилатциклазной системы передачи сигнала. Аденилатциклаза обнаружена во всех типах клеток.

Рис. 5-36. Образование циклического аденозинмонофосфата (цАМФ).
Фермент относят к группе интегральных белков клеточной мембраны, он имеет 12 трансмембранных доменов. Внеклеточные фрагменты аденилатциклазы гликозилированы. Цитоплазматические домены аденилатциклазы имеют два каталитических центра, ответственных за образование цАМФ - вторичного по средника, участвующего в регуляции активности фермента протеинкиназы А.
На активность аденилатциклазы оказывают влияние как внеклеточные, так и внутриклеточные регуляторы. Внеклеточные регулятор (гормоны, эйкозаноиды, биогенные амины) осуществляют регуляцию через специфические рецепторы, которые с помощью α-субъединиц G-белков передают сигналы на аденилатциклазу. αs - Субъединица (стимулирующая) при взаимодействии с аденилатциклазой активирует фермент, αi- Субъединица (ингибирующая) ингибирует фермент. В свою очередь, аденилатциклаза стимурирует проявление ГТФ- фосфотазной активности α- субъединиц. В результате дефосфорилирования ГТФ образуются субъединицы а s -ГДФ и а i -ГДФ, не комплементарные аденилатциклазе.
Из 8 изученных изоформ аденилатциклазы 4 - Са 2+ -зависимые (активируются Са 2+). Регуляция аденилатциклазы внутриклеточным кальцием позволяет клетке интегрировать активность двух основных вторичных посредников цАМФ и Са 2+ .
Д. фосфолипазы
Фосфолипазы - ферменты класса гидролаз, катализирующие катаболизм глицерофосфолипидов. Различают фосфолипазы секреторные, входящие в состав панкреатического сока, и клеточные фосфолипазы. Клеточные фосфолипазы А 1 , А 2 , D, С различаются по специфичности к отщепляемой группе. Все фосфолипазы - кальций зависимые ферменты (рис. 5-37).

Рис. 5-37. Действие фосфолипаз.
Фосфолипаза С - фермент, гидролизующий фосфоэфирную связь в глицерофосфолипидах. В клетках человека идентифицировано 10 изоформ фосфолипазы С, различающихся по молекулярной массе, локализации, способу регуляции, субстратной специфичности. В структуре всех изоформ фосфолипазы С отсутствуют гидрофобные домены, которые могли бы обеспечить их взаимодействие с мембраной. Однако некоторые формы фосфолипазы С связаны с помощью гидрофобного «якоря» - ацильного остатка миристиновой кислоты или за счёт взаимодействия с поверхностью бислоя. Каталитическая активность всех изоформ фосфолипазы С зависит от ионов кальция.
Большинство фосфолипаз С специфично в отношении фосфатидилинозитолов и практически не гидролизует другие типы фосфолипидов. Активный фермент может гидролизовать до 50% от общего количества фосфатидилинозитолов клеточной мембраны. При гидролизе фосфатидилинозитол-4,5-бисфосфата (ФИФ 2) образуются продукты диацилглицерол (ДАГ) и инозитол-1,4,5-трифосфат (ИФ 3), служащие вторичными посредниками в трансмембранной передаче сигнала по инозитолфосфатному пути.
Е. протеинкиназы
Все полярные сигнальные молекулы, действующие на клетку-мишень через мембранные рецепторы, осуществляют свою биологическую функцию путём фосфорилирования специфических белков и ферментов, регулирующих метаболизм в клетке. Фосфорилирование изменяет (увеличивает или уменьшает) их активность. Катализируют фосфорилирование белков (протеинов) протеинкиназы по аминокислотным остаткам серина, треонина, тирозина. Протеинкиназы могут быть субъединицей мембранного рецептора, например тирозиновая протеинкиназа рецептора инсулина, активность которой регулируется гормоном. Другая группа - протеинкиназы, регулируемые вторичными вестниками гормонального сигнала (цАМФ, цГМФ, Са 2+ , ДАГ), например протеинкиназа А, протеинкиназа С, протеинкиназа G, кальмодулинзависимые протеинкиназы и др.
Протеинкиназы А
Протеинкиназы А (цАМФ-стимулируемые) участвуют в аденилатциклазной системе передачи сигнала. Протеинкиназа А состоит из 4 субъединиц R 2 С 2 - двух регуляторных субъединиц (R 2) и двух каталитических (С 2) (см. рис. 5-41). Комплекс R 2 С 2 не обладает ферментативной активностью.
Комплекс R 2 С 2 разными способами прикрепляется к мембране. Некоторые формы протеинкиназы А «заякориваются» с помощью алифатического остатка миристиновой кислоты каталитических субъединиц. Во многих тканях протеинкиназа А связана с «заякоренным» белком АКАР s (от англ. сАМР dependent protein kinase anchoring proteins). АКАР s имеет центр связывания для регуляторных субъединиц протеинкиназы А. С помощью белка АКАР s протеинкиназа А связывается с мембраной в области локализации ферментов, катализирующих образование цАМФ (аденилатциклаза) или его гидролиз (фосфодиэстераза), а также белков, в регуляции активности которых фермент принимает участие, например потенциалзависимые Са 2+ -каналы.
Регуляторные субъединицы протеинкиназы А имеют специфические центры для связывания цАМФ. Присоединение цАМФ к регуляторным, субъединицам приводит к изменению конформации последних и снижению сродства к каталитическим субъединицам С, происходит диссоциация по схеме:
цАМФ 4 + R 2 С 2 -> цАМФ 4 R 2 + С + С
Субъединицы С представляют собой активную форму протеинкиназы А, которая катализирует реакции фосфорилирования по серину и треонину. Каталитические субъединицы С у разных типов протеинкиназ а не идентичны, они различаются прежде всего специфичностью в отношении белков-субстратов.
Протеинкиназы С
Протеинкиназы С участвуют в инозитолфосфатной системе передачи сигнала. Фермент состоит из двух функционально различных доменов - регуляторного и каталитического. Регуляторный домен содержит 2 структуры («цинковые пальцы»), образованные фрагментами пептидной цепи, богатыми цистеином, и содержащими 2 иона цинка (см. раздел 1). «Цинковые пальцы» участвуют в связывании диацилглицерола. Другой фрагмент регуляторного домена имеет высокое сродство к Са 2+ . Повышение концентрации кальция в цитозоле увеличивает сродство протеинкиназы С к фосфатидилсерину мембраны. Транслокация протеинкиназы С к мембране позволяет ферменту связаться с ДАГ, который ещё больше повышает сродство протеинкиназы С к ионам кальция (рис. 5-38). Наиболее распространённые изоформы протеинкиназы С активируются Са 2+ , диацилглицеролом и фосфатидилсерином.

5-38. Регуляция активности протеинкиназы С (ПКС). ФС - фосфатидилсерин; ДАГ - диаципглицерол.
Каталитический домен имеет центр, связывающий АТФ и белок-субстрат. Активная фермента протеинкиназы С фосфорилирует по остаткам серина и треонина. Снижение концентрации ионов кальция в клетке нарушает связь протеинкиназы С с фосфатидилсерином и диацилглицеролом, фермент переходит в неактивную форму и отделяется от мембраны.
3. Протеинкиназы G
В отличие от протеинкиназы А, протеинкиназа G присутствует не во всех тканях, ее обнаруживают в лёгких, мозжечке, гладких мышцах и тромбоцитах. Изоформы протеинкиназы G могут быть связаны с мембраной или находиться цитоплазме. Растворимая протеинкиназа С состоит из двух идентичных субъединиц, каждая из которых имеет два центра для связывания цГМФ. Приединение цГМФ к регуляторным центрам вызывает конформационные изменения субъединиц и повышает каталитическую активность фермента (рис. 5-39). Протеинкиназа G, подобно протеинкиназе А и С, специфична в отношении определённых белковых субстратов, которые она фосфорилирует по остаткам серина и треонина.

Рис. 5-39. Регуляция активности протеинкиназы G (ПКG).
Ж фосфодиэстеразы
Фосфодиэстеразы - ферменты, катализирующие превращение цАМФ (рис. 5-40) или цГМФ в неактивные метаболиты АМФ или ГМФ. Фосфодиэстеразы, снижая концентрации вторичных посредников, разрывают цепь превращений, вызванных активатором рецептора.

Рис.5-40. Превращение цАМФ в АМФ.
Фосфодиэстеразы присутствуют в клетках тканей в 2 формах: в форме растворимого белка и мембранносвязанного. Формы фермента, связанные с мембраной, в разных тканях составляют 5-40%. В одной и той же ткани могут присутствовать разные формы фосфодиэстеразы, различающиеся по сродству к субстратам, молекулярному весу, заряду, регуляторным свойствам и локализации в клетке.
Фосфодиэстеразы циклических нуклеотидов не обладают абсолютной специфичностью, поэтому, как правило, одна и та же форма фермента способна гидролизовать как цАМФ, так и цГМФ. Однако скорости гидролиза этих двух нуклеотидов под действием одной и той же фосфодиэстеразы могут значительно различаться. Это зависит от того, какая фосфодиэстераза присутствует в клетке - более специфичная в отношении цАМФ или более специфичная к цГМФ, от соотношения концентраций цАМФ и цГМФ в клетке и от действия регуляторов фосфодиэстеразы.
В большинстве тканей присутствует фосфодиэстераза-1, более специфичная к цАМФ, активируемая Са 2+ , комплексом 4 Са 2+ -кальмодулин и цГМФ.
ТРАНСМЕМБРАННАЯ ПЕРЕДАЧА СИГНАЛА. Важное свойство мембран - способность воспринимать и передавать внутрь клетки сигналы из внешней среды. "Узнавание" сигнальных молекул осуществляется с помощью белков-рецепторов, встроенных в клеточную мембрану клеток-мишеней или находящихся в клетке.
Если сигнал воспринимается мембранными рецепторами, то схему передачи информации можно представить так:
взаимодействие рецептора с сигнальной молекулой (первичным посредником);
активация мембранного фермента, ответственного за образование вторичного посредника;
образование вторичного посредника цАМФ, цГМФ, ИФ3, ДАТ или Са 2+ ;
активация посредниками специфических белков, в основном протеинкиназ, которые, в свою очередь, фосфорилируя ферменты, оказывают влияние на активность внутриклеточных процессов.
Существует несколько механизмов трансмембранной передачи информации: с использованием аденилатциклазной системы, инозитолфосфатной системы, каталитических рецепторов, цитоплазматических или ядерных рецепторов.
Cтруктурно-функциональная организация G-белков
G-белки (ГТФ-связывающие белки) - универсальные посредники при передаче сигналов от рецепторов к ферментам клеточной мембраны, катализирующим образование вторичных посредников гормонального сигнала. G-белки - олигомеры, состоящие из α, β и γ-субъединиц.
Каждая α-субъединица в составе G-белка имеет специфические центры:
связывания ГТФ или ГДФ;
взаимодействия с рецептором;
связывания с βγ-субъединицами;
фосфорилирования под действием протеинкиназы С;
взаимодействия с ферментом аденилатциклазой или фосфолипазой С.
В структуре G-белков отсутствуют α-спиральные, пронизывающие мембрану домены. G-белки относят к группе "заякоренных" белков.
Регуляция активности G-белков
Различают неактивную форму G-белка - комплекс αβγ-ГДФ и активированную форму αβγ-ГТФ. Активация G-белка происходит при взаимодействии с комплексом активатор-рецептор, изменение конформации G-белка снижает сродство α-субъединицы к молекуле ГДФ и увеличивает к ГТФ. Замена ГДФ на ГТФ в активном центре G-белка нарушает комплементарность между α-ГТФ и βγ-субъединицами. Рецептор, связанный с сигнальной молекулой, может активировать большое количество молекул G-белка, таким образом обеспечивая усиление внеклеточного сигнала на этом этапе.
Активированная α-субъединица G-белка (α-ГТФ) взаимодействует со специфическим белком клеточной мембраны и изменяет его активность. Такими белками могут быть ферменты аденилатциклаза, фосфолипаза С, фосфоди-эстераза цГМФ, Nа+-каналы, К+-каналы.
Рис. 5-35. Цикл функционирования G-белка. R s - рецептор; Г - гормон; АЦ - аденилатциклаза.
Следующий этап цикла функционирования G-белка - дефосфорилирование ГТФ, связанного с α-субъединицей, причём фермент, катализирующий эту реакцию, - сама α-субъединица.
Дефосфорилирование приводит к образованию комплекса α-ГДФ, который не комплементарен специфическому белку мембраны (например, аденилатциклазе), но имеет высокое сродство к ру-протомерам. G-белок возвращается к неактивной форме - αβγ-ГДФ. При последующей активации рецептора и замене молекулы ГДФ на ГТФ цикл повторяется снова. Таким образом, α-субъединицы G-белков совершают челночное движение, перенося стимулирующий или ингибирующий сигнал от рецептора, который активирован первичным посредником (например, гормоном), на фермент, катализирующий образование вторичного посредника.
Некоторые формы протеинкиназ могут фосфорилировать α-субъединицы G-белков. Фосфорилированная α-субъединица не комплементарна специфическому белку мембраны, например аденилатциклазе или фосфолипазе С, поэтому не может участвовать в передаче сигнала.
Аденилатциклаза
Фермент аденилатциклаза, катализирующий превращение АТФ в цАМФ, - ключевой фермент аденилатциклазной системы передачи сигнала. Фермент относят к группе интегральных белков клеточной мембраны, он имеет 12 трансмембранных доменов. Внеклеточные фрагменты аденилатциклазы гликозилированы. Цитоплазматические домены аденилатциклазы имеют два каталитических центра, ответственных за образование цАМФ - вторичного посредника, участвующего в регуляции активности фермента протеинкиназы А.
На активность аденилатциклазы оказывают влияние как внеклеточные, так и внутриклеточные регуляторы. Внеклеточные регуляторы (гормоны, эйкозаноиды, биогенные амины) осуществляют регуляцию через специфические рецепторы, которые с помощью α-субъединиц G-белков передают сигналы на аденилатциклазу. α s -Субъединица (стимулирующая) при взаимодействии с аденилатциклазой активирует фермент, α-субъединица (ингибирующая) ингибирует фермент. Из 8 изученных изоформ аденилатциклазы 4 - Са 2+ -зависимые (активируются Са 2+). Регуляция аденилатциклазы внутриклеточным кальцием позволяет клетке интегрировать активность двух основных вторичных посредников цАМФ и Са 2+ .
Аденилатциклазная система
При участии аденилатциклазной системы реализуются эффекты сотни различных по своей природе сигнальных молекул - гормонов, нейромедиаторов, эйкозаноидов.
Функционирование системы трансмембранной передачи сигналов обеспечивают белки: Rs-рецептор сигнальной молекулы, которая активирует аденилатциклазу, и R i -рецептор сигнальной молекулы, которая ингибирует аденилатциклазу; G s -стимулирующий и G j -ингибирующий аденилатциклазу белки; ферменты аденилатциклаза (АЦ) и протеинкиназа А (ПКА).
Последовательность событий, приводящих к активации аденилатциклазы:
связывание активатора аденилатциклазной системы, например гормона (Г) с рецептором (R s), приводит к изменению конфор-мации рецептора и увеличению его сродства к G s -белку. В результате образуется комплекс [Г][R][О-ГДФ];
присоединение [Г][R] к G-ГДФ снижает сродство α-субъединицы G s -белка к ГДФ и увеличивает сродство к ГТФ. ГДФ замещается на ГТФ;
это вызывает диссоциацию комплекса. Отделившаяся субъединица α, связанная с молекулой ГТФ, обладает сродством к адени-латциклазе:
[Г][R] → [Г][R] + α-ГТФ + βγ;
взаимодействие α-субъединицы с аденилатциклазой приводит к изменению конфор-мации
фермента и его активации, увеличивается скорость образования цАМФ из АТФ;
конформационные изменения в комплексе [α-ГТФ][АЦ] стимулируют повышение ГТФ-фосфатазной активности α-субъединицы. Протекает реакция дефосфорилирования ГТФ, и один из продуктов реакции - неорганический фосфат (P i) отделяется от α-субъединицы, а комплекс [α-ГДФ] сохраняется; скорость гидролиза определяет время проведения сигнала;
образование в активном центре α-субъединицы молекулы ГДФ снижает его сродство к аденилатциклазе, но увеличивает сродство к βγ-субъединицам. G s -белок возвращается к неактивной форме;
если рецептор связан с активатором, например гормоном, цикл функционирования G s белка повторяется.

Аденилатциклазная система
Цикл активации G-белка под действием G-белок-связанного рецептора.
G-белки - это семейство белков, относящихся к ГТФазам и функционирующих в качестве вторичных посредников во внутриклеточных сигнальных каскадах. G-белки названы так, поскольку в своём сигнальном механизме они используют замену GDP на GTP как молекулярный функциональный «выключатель» для регулировки клеточных процессов.
История
G-белки были обнаружены и исследованы Альфредом Гилманом и Мартином Родбеллом, которые получили за это открытие Нобелевскую премию по физиологии и медицине 1994 года.
Типы G-белков
G-белки делятся на две основных группы - гетеротримерные и «малые». Гетеротримерные G-белки - это белки с четвертичной структурой, состоящие из трёх субъединиц: альфа, бета и гамма. Малые G-белки - это белки из одной полипептидной цепи, они имеют молекулярную массу 20-25 кДа и относятся к суперсемейству Ras малых ГТФаз. Их единственная полипептидная цепь гомологична α-субъединице гетеротримерных G-белков. Обе группы G-белков участвуют во внутриклеточной сигнализации.
Гетеротримерные G-белки
У всех гетеротримерных G-белков сходный механизм активации: они активируются при взаимодействии со специфическими рецепторами, сопряженными с G-белками, при этом обменивая ГДФ на ГТФ и распадаясь на α- и βγ-субъединицы. α-субъединица, связанная с ГТФ, воздействует на следующее звено в цепи передачи сигнала. βγ-субъединица также может вызывать собственные эффекты. Инактивация G-белков происходит в результате медленного гидролиза ГТФ до ГДФ α-субъединицей, после чего происходит реассоциация субъединиц.



